
CYTOSQUELETTE
– Regroupé dans les cellules eucaryotes un ensemble de polymères fibreux (cytosoliques et nucléaires) et de protéines associées ; joue le rôle d’un véritable « squelette cellulaire » en déterminant la forme des cellules, des organites, du noyau et en participant à la polarité des cellules ; joue également le rôle d’une musculature cellulaire, responsable des mouvements des cellules elles-mêmes ou des composants cellulaires à l’intérieur des cellules.
– Le cytosquelette est constitué de trois classes de filaments non spécifiques et ubiquitaires :
→ Les microfilaments d’actine = MFA ( ∅= 8 nm).
→ Les filaments intermédiaires = FI ( ∅= 10 nm).
→ Les microtubules = MT ( ∅= 25 nm).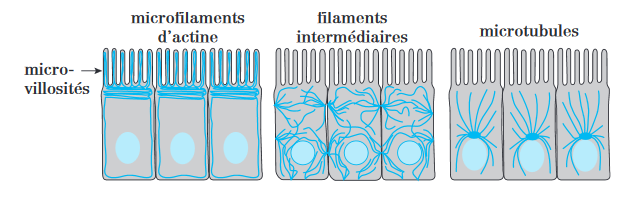
– Deux types de monomères protéiques sont à la base des polymères fibreux du cytosquelette :
→ monomères globulaires pour les MFA et les MT.
→ monomères fibreux pour les FI.
– Les éléments du cytosquelette existent sous trois formes en équilibre dans la cellule :
→ monomères libres néosynthétisés ou issus de la dépolymérisation ;
→ polymères instables car leur fréquence de polymérisation/dépolymérisation est élevée ;
→ polymères stabilisés par des interactions avec des protéines associées.
– Les éléments du cytosquelette se localisent dans les trois compartiments cellulaires suivants :
→ le cytosol.
→ le nucléoplasme (en particulier, les lamines qui sont des FI).
→ la périphérie de la cellule, sous la membrane plasmique, où ils forment le cortex cellulaire.
MICROFILAMENTS D’ACTINE
– Microfilaments d’actines ou actine f (fibrilaire) sont des polymères d’actine G qui constitue 2 protofilaments suivant une disposition monocaténaire en hélice. Ils sont ubiquitaires chez les eucaryotes.
– L’actine est l’une des protéines cellulaires les plus abondantes : 1 à 5 % de l’ensemble des protéines dans les cellules non musculaires (sous forme d’actine β et γ) et 20 % dans les cellules musculaires (sous forme d’actines α).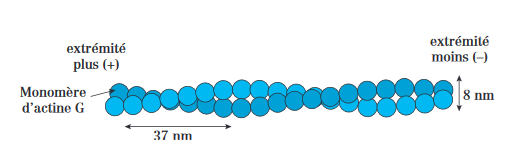
1-BIOSYNTHESE :
– Le complexe protéique ARP2/3 se fixe côté (–) de l’actine et favorise la formation d’une amorce constituée de trimères d’actine G ( noyau) qui s’assemblent en filaments polarisés par une élongation/polymérisation.
– Dans l’extremité + la polymérisation est rapide alors que la polymérisation est plus lente dans l’extrémité — .
– Il existe des drogues qui perturbent la polymérisation tel que les cytochalasines,
*Remarque : molécule globulaire constituée par deux lobes et possède deux domaines séparés par un sillon, au fond de celui-ci est localisé les sites de liaisons pour Mg 2+ et de l’ATP ou L’ADP.
2-PROTÉINES ASSOCIÉS (selon leurs fonctions)
2.1-RÉGULATION :
– Profiline——————> favorise la polymérisation
– Caldesmon et CAP——> empêche la dépolymérisation (stabilisation)
– Thymosine—————> inhibe (block) la polymérisation
– ARP2/3——————-> catalise la nucléation
2.2-FRAGMENTATION ; ex: la gélsonine
2.3-ANCRAGE MENBRANAIRES :
– miosine I———> attachent les microvillosités à la mb plasmique
– spectrine———> sous la mb plasmique et attache la micro filaments d’actine à la mb plasmique
2.4-CONTRACTION MUSCULAIRE : La miosine II constituée de 2 têtes bipolaire à activité ATPasique en présence de Ca++-. Elles se phosphorilisent et s’organisent en myofilaments liés au MF d’actine formant un complexe actomyosine.
2.5-MOUVEMENTS DE VESICULES ET D’ORGANITES : Qui se fait le long des microfilaments par la miosine I jusqu’à l’extrémité (+).
2.6-ORGANISATION :
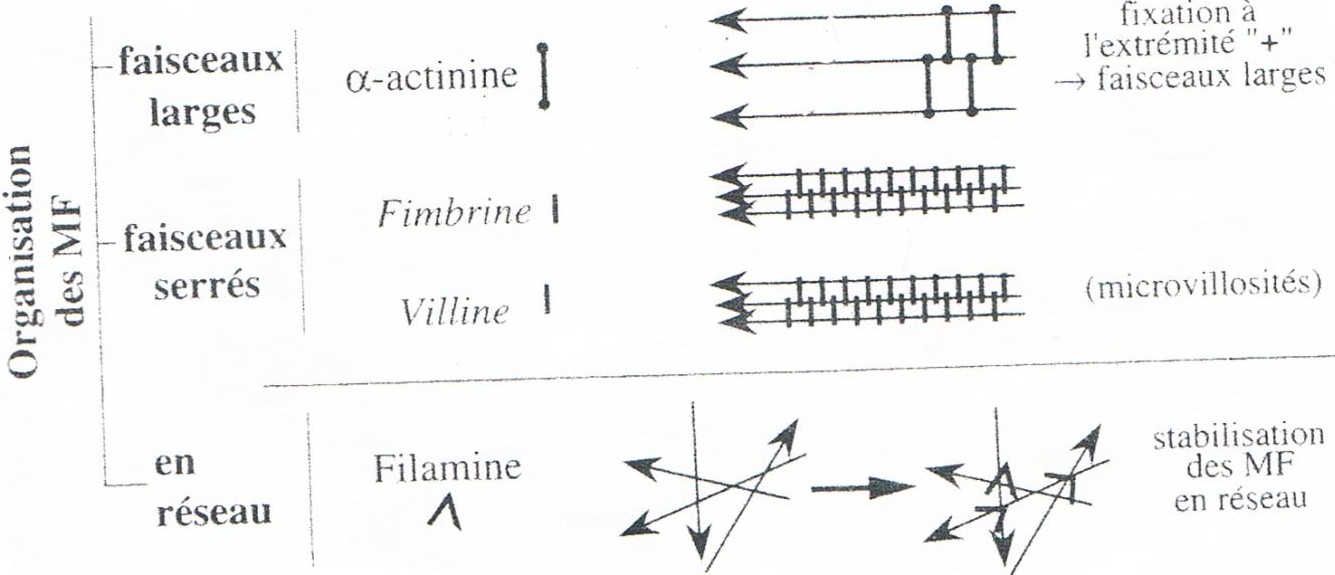
R ! Les faisceaux serrés au niveau microvillosités des entérocytes et les faisceaux large au niveau des fibres musculaires striées.
3-FONCTIONS :
- – La forme et le maintien de la polarité des cellules (microvillosités apicales).
- Les mouvements intracellulaires des organites: mouvements de cyclose (cellule végétale).
- Les mouvements cellulaires : déplacement de l’amibe, des leucocytes (pseudopodes).
- La cytodiérèse (au cours de la division cellulaire)
- – La formation des jonctions cellulaires.
LES MICROTUBLUES
1-ULTRASTRUCTURE ET BIOGENÈSE :
– Polymères instables et polarisés, forme de tube creux de diamètre plus grand que celui des MF d’actine (25 nm). Ils sont constitués de deux types de protéines globulaires, les tubulines alpha (α) et béta (β) qui s’associent en dimères. La polymérisation se fait en présence de GTP et Mg ++. Les dimères se polymérisent en protofilaments et l’association de 13 protofilaments forme un MT.
– Les MT présentent deux extrémités qui s’allongent à des vitesses différentes. L’extrémité (+) à polymérisation rapide vers l’extérieur, l’extrémité (-) à polymérisation plus lente vers le centre cellulaire.
– La polymérisation est liée a l’interaction de la tubuline β avec le GTP et la dépolymérisation est précédée par l’hydrolyse de GTP en GDP, et ceci dans les centres organisateurs (MTOC) dont: les kinétochores (divisions cellulaires), le corpuscule basal ou le cinétosome (cils et flagelles), et enfin la matière péricentriolaire amorphe (cellules animales) ou masse amorphe (cellules végétales) pour la polymérisation des autres MT.
– Certaines drogues perturbent la polymérisation/dépolymérisation des MT et bloquent la division cellulaire :
- La colchicine et la vinblastine bloquent la polymérisation des MT en séquestrant les monomères libres de tubuline. Comme elles ne bloquent pas la dépolymérisation, les MT raccourcissent.
- Le taxol stabilise les microtubules polymérisés et inhibe la dépolymérisation des microtubules.
2-PROTEINES ASSOCIÉES (MAP) :
– structurales: MAP1, MAP2 et TAU qui n’est présente que dans les neurones.
– motrices: deplacements orientés de molécules, de vésicules ou d’organites le long des microtubules.
• Les kinésines transportent vers l’extrémité (+), distale, des microtubules (transport antérograde).
• Les dynéines transportent vers l’extrémité (–), proximale, des microtubules (transport rétrograde).
– Ces protéines sont organisés en trois domaines: deux têtes identiques se fixant sur les microtubules et possédant une activité ATPasique, une tige et une queue ou se fixe le matériel à transporter via des protéines intermédiaire ( kinectine et dynactine).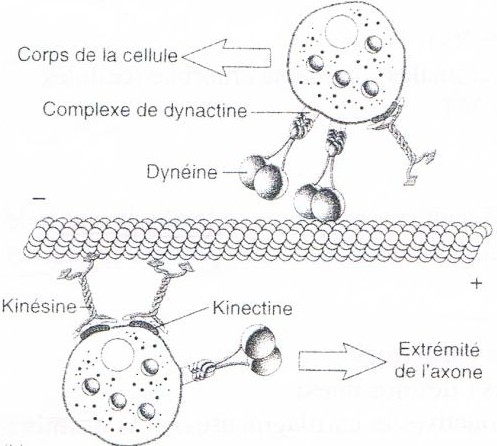
3-VARIÉTÉS DE MT :
3.1-STABLE: conservés quelque soit le fixateur et la température de fixation et constituent les éléments cellulaires permanents.
3.1.1-LES CENTRIOLES :
• Le centrosome est composé de deux centrioles, disposés perpendiculairement l’un à l’autre, baignant dans le matériel péricentriolaire. Chaque centriole est un cylindre creux constitué de 9 triplets de microtubules réunis par des liaison de nexine. Le microtubule le plus interne de chaque triplet, appelé microtubule A, est complet (13 protofilaments). Les microtubules distaux (B et C) sont incomplets.
• Le microtubule C est relié au A par des liaisons transversales. On note l’absence de centrioles dans la cellule végétale. Le centriole, en se divisant, peut fournir des corpuscules basaux.
3.1.2-LES CILS ET FLAGELLES :
• des expansions de la membrane plasmique contenant un squelette organisé de microtubules appelé axonème, et ancrées dans le cytosol par un corpuscule basal. L’axonème est le prolongement du corpuscule basal, il est composé de 9 doublets de microtubules périphériques et de 2 microtubules centraux reliés entre eux entourés par un manchon de MAP.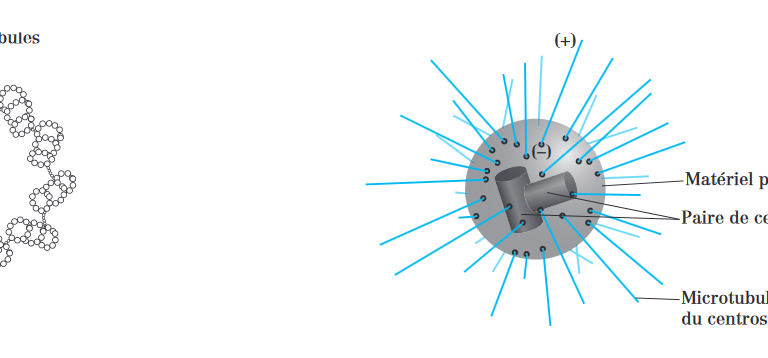
3.2-LABILES:
• conservés par des fixateurs aldéhydiques (glutaraldéhvde) et à température supérieure à 4° C, (fuseau mitotique au moment de la division cellulaire) contribuent au mouvements des chromosomes grâce a la poly et la dépolymérisation.
4-FONCTIONS :
• le maintien de la forme de la cellule (polarité).
• le déplacement des chromosomes (mitose et meïose).
• le transport des vésicules d’endocytose et d’exocytose.
• le déplacement des organites intra cellulaires.
• le flux axonal c.a.d le mouvement des vésicules synaptiques vers la synapse.
• le mouvement de cellules isolées (paramécie, spermatozoïde, .) par l’intermédiaire des cils et flagelles.
FILAMENTS INTERMEDIAIRES
– Deux monomères de même orientation s’associent par leur domaine central (interactions hydrophobes) pour donner un dimère torsadé (association parallèle) puis ils s’associent avec un décalage pour former un tétramère (association anti-parallèle) et l’association bout à bout de plusieurs tétramères constitue un protofilament. 8 protofilaments s’associent pour former un FI de forme cylindrique de 8 à 10 nm de large diamètre intermédiaire entre celui des microfilaments d’actine et les microtubules, d’où son nom.
– Les FI sont présents dans le hyaloplasme (tout autour du noyau “corbeille péri-nucléaire” sauf au ports nucléaires) et le nucléoplasme, Ce sont des polymères stables formés de protéines fibreuses qui sont :
| types | familles | localisation |
| 1 |
Kératines acides |
situés au niveau des cellules épithéliailes (desmosomes et hémi-desmosomes) et les dérivés épidermiques (ongles, cheveux, poils…). |
| 2 |
Kératines basiques |
|
| 3 | Desmine |
situés au niveau des muscles lisses et striés, ils relient les myofilaments entre eux et à la mb plasmique. |
| GFAP |
trouvée dans deux types de cellules gliales (les astrocytes et les cellules de Schwann). |
|
| Vimentine |
situés au niveau des tissus qui dérivent du mésenchyme (muscles lisses, vaisseaux sanguins), fibro-blastes, cellules endothéliales… |
|
| 4 | neurofilaments | situés au niveau des neurones, où ils s’associent aux microtubules. |
| 5 |
Lamines (A, B et C) |
situés au niveau du noyau où ils forment le réseau péri-nucléaire. |
| 6 | Nestine | Ce localisent dans les Cellules neuroépithéliales (souches) |
– Il existe des pros associés aux filaments intermédiaires à rôle organisateur du CS :
• Filaggrine : Présente dans les kératinocytes, synthétisées dans la couche granuleuse de l’épiderme.
• Plakine : Responsable des liaisons des Fi avec les autres constituants du cytosquelette.
• Plectine et BPAG1 : Présente dans les hémidesmosomes (cohésion cellulaire).
1-FONCTIONS : Elle concernent principalement l’architecture cellulaire et tissulaire et elles dépendent essentiellement du type de filament intermédiaire :
• Dans les épithéliums, les cytokératines relient les cellules entre elles par l’intermédiaire des desmosomes. Elles assurent ainsi leur cohésion et leur stabilité mécanique ;
• Dans les cellules nerveuses, les neurofilaments assurent la continuité et l’élasticité des neurones ;
• Dans les noyaux, les lamines assurent la stabilité de l’enveloppe nucléaire interne et son interaction avec la chromatine.
*Fonctions du cytosquelette = fonctions des (MFA+FI+MT)