
SYSTEME NERVEUX AUTONOME
A-Introduction :
Le système nerveux autonome est une partie du système nerveux périphérique, à l’instar du système nerveux somatique. Il contribue au maintien de l’homéostasie en régulant de nombreux processus physiologiques et métaboliques.
Il innerve notamment les viscères, les muscles lisses, le cœur, les vaisseaux… Son action est dite involontaire car elle ne nécessite pas l’intervention de la volonté. C’est un système effecteur..
Il possède deux subdivisions : le système nerveux parasympathique et le système nerveux orthosympathique (aussi appelé système nerveux sympathique).
La plupart des organes reçoivent une double innervation, mis à part les glandes sudoripares, les muscles pilomoteurs et quelques vaisseaux sanguins strictement orthosympathiques.
Il fonctionne de façon réflexe : récepteur périphérique à fibres afférentes sensorielles à centre végétatif à fibres efférentes motrices à organes efférents.
B-Anatomie fonctionnelle :
Généralités :
L’innervation motrice des viscères comporte deux types de cellules :
- Le neurone préganglionnaire : c’est une fibre faiblement myélinisée de type B dont le soma est situé dans le centre végétatif, et dont l’axone chemine jusqu’au ganglion végétatif.
- Le neurone postganglionnaire : c’est une fibre amyélinique de type C, dont le soma se trouve au niveau du ganglion végétatif et dont l’axone chemine jusqu’aux viscères.
- Les neurones pré et postganglionnaires forment une synapse dans le ganglion végétatif.
Système orthosympathique :
C’est un système exclusivement d’origine spinale thoracolombaire. Il s’étend des segments D2 à L3. Son circuit est composé de deux types de neurones :
- Neurones sympathiques préganglionnaires :
Leurs somas se situent dans le noyau intermédiolatéral de la couche VII de Rexed. Ils envoient des fibres efférentes par la corne ventrale de la moelle épinière puis vers le nerf rachidien. Les fibres abandonnent ensuite le nerf rachidien pour rejoindre la chaine sympathique paravertébrale en constituant un rameau communicant blanc. Il a ensuite trois destinées possibles :
- Une synapse avec un neurone sympathique postganglionnaire du ganglion paravertébral du même étage de la chaîne sympathique.
- Une synapse avec un neurone sympathique postganglionnaire du ganglion paravertébral d’un étage plus crânial ou plus caudal de la chaine sympathique.
- Emprunte le nerf sympathique pour ensuite former une synapse avec un neurone postganglionnaire dans un ganglion prévertébral à distance de la chaîne sympathique paravertébrale.
- Neurones sympathiques postganglionnaires :
Leurs somas se situent dans les ganglions de la chaine sympathique paravertébrale ou dans les ganglions prévertébraux (à distance).
- Ceux qui se situent dans la chaine sympathique paravertébrale rejoignent le nerf rachidien en empruntant un rameau communicant gris.
- Ceux qui se situent dans le ganglion prévertébral empruntent le nerf sympathique pour aller innerver le viscère ciblé.
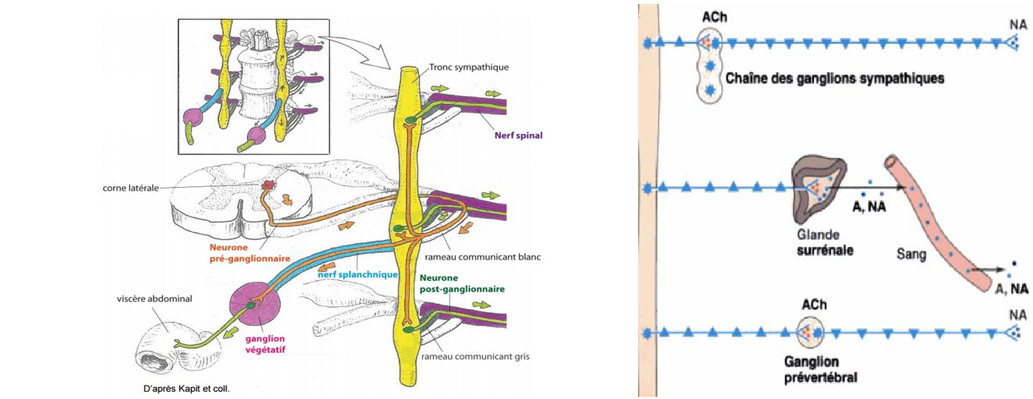
- Glande médullosurrénale :
La médullosurrénale constitue un cas particulier équivalent au « neurone postganglionnaire ». La fibre préganglionnaire se termine directement sur les cellules chromaffines de la glande, qui sécrètent des monoamines (adrénaline et noradrénaline). - Le ganglion sympathique constitue un relais de transmission de l’information. Au niveau de ce relais, un neurone préganglionnaire peut faire synapse avec plusieurs neurones
- On dit qu’il y a dispersion axonale. Le ratio est d’1 fibre pré pour 20 à 30 fibres
- Les effecteurs sont distribués dans tout le corps, même au niveau céphalique.
- L’axone du neurone préganglionnaire est court dû à la proximité des ganglions
Système parasympathique :
Les neurones préganglionnaires du système parasympathique ont une double origine : crâniale (du tronc cérébral) ou sacrale. Leurs axones sont plus longs que ceux du système orthosympathique étant-donné que les ganglions parasympathiques sont situés près des organes cibles (parfois dans leur paroi même).
Contingent céphalique :
Représentés par les 3ème, 7ème, 9ème, et 10ème paires de nerfs crâniens, prenant naissance respectivement au niveau des noyaux du mésencéphale pour ensuite cheminer dans les nerfs crâniens.
- Noyau d’Edinger-Westphal du III (nerf oculomoteur) : destinés aux sphincters pupillaires et aux muscles ciliaires de l’œil.
- Noyau salivaire supérieur du VII (nerf facial) : destinés aux glandes lacrymales, nasales, sublinguales et sous-maxillaires.
- Noyau salivaire inférieur du IX (nerf glossopharyngien) : destinés aux glandes
- Noyau dorsal du X (nerf vague) : destinés au cœur, aux bronches, à l’œsophage, à l’estomac, à l’intestin proximal, au foie, à la vésicule biliaire, au pancréas et à la partie supérieure des uretères.
75% des fibres parasympathiques cheminent dans le nerf vague.
Contingent sacral :
Ses fibres sont issues des racines S2 à S3 (pouvant aller de S1 à S4). Elles sortent par les racines ventrales sacrales pour se terminer sur la paroi des viscères.
Elles innervent notamment la partie distale du côlon (descendant et rectum), la partie basse de l’urètre et les tissus érectiles des organes génitaux.
Différences structurales entre les deux systèmes :
| Sympathique Parasympathique | ||
| Origine de la fibre présynaptique Localisation du ganglion
Longueur de la fibre présynaptique |
Thoraco-lombaire | Crânio-sacrale |
| Près du SNC | Près de l’organe cible | |
|
Courte |
Longue |
|
C-Transmission synaptique dans le SNA :
Généralités :
D’une manière générale, les neurotransmetteurs libérés par le neurone présynaptique agissent sur des récepteurs spécifiques de la membrane postsynaptique.
Il existe deux types de récepteurs :
- Les récepteurs ionotropes : induisent des modifications de la perméabilité membranaire à certains ions, modifiant ainsi le potentiel de membrane de la cellule cible.
- Les récepteurs métabotropes : activent certains systèmes enzymatiques (adénylate cyclase) afin de moduler les activités de la cellule.
Synapses au niveau du ganglion végétatif :
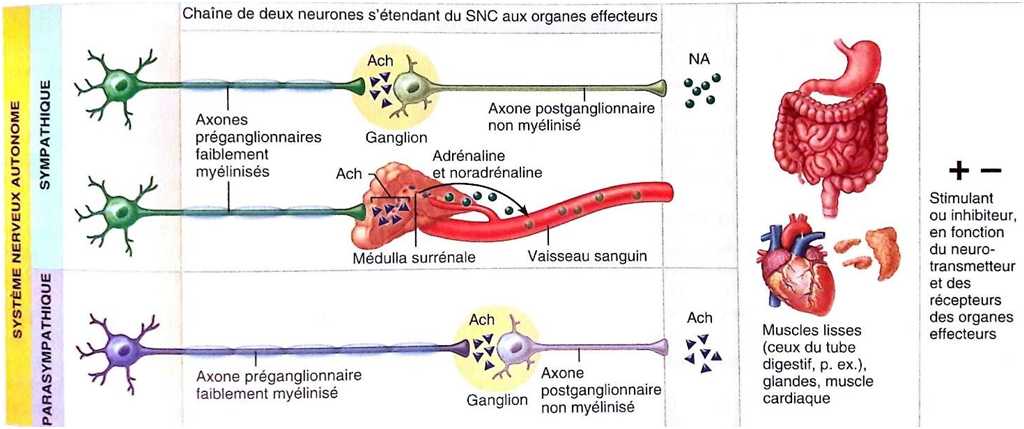
Dans le SNA, tous les neurones préganglionnaires qu’ils soient sympathiques ou parasympathiques, sont de type cholinergique. Cependant, les neurones préganglionnaires sympathiques peuvent parfois sécréter des neuropeptide (enképhalines, LH-RH, substance P, somatostatine…).
Les boutons synaptiques des neurones préganglionnaires sympathiques et parasympathiques libèrent de l’acétylcholine qui va se fixer sur ses récepteurs spécifiques de type nicotinique ou muscarinique au niveau de la membrane post-synaptique du neurone postganglionnaire.
La stimulation du neurone présynaptique induit une réponse postsynaptique sous forme de potentiel triphasique.
L’on distingue trois réponses postsynaptiques possibles :
- Une dépolarisation PPSE : due à l’excitation par les récepteurs cholinergiques nicotiniques.
- Une hyperpolarisation PPSI : due à l’inhibition par des interneurones adrénergiques.
- Une dépolarisation PPSE : en rapport avec une transmission muscarinique tardive, bloquée par l’atropine.
Remarque :
- Les agonistes nicotiniques excitent simultanément les systèmes sympathique et parasympathique au niveau des ganglions végétatifs.
- La nicotine ne peut pas stimuler directement les organes effecteurs car leurs membranes possèdent des récepteurs de type muscariniques.
- L’acétylcholine quant à elle active aussi bien les récepteurs nicotiniques que muscariniques.
- Il existe des agents bloquants nommés ganglioplégiques qui empêchent la transmission synaptique au niveau des ganglions végétatifs sympathiques et parasympathiques, tels que le TEA, l’héxaméthonium, le pentolinium…
Synapses au niveau de l’organe effecteur :
-
-
- Les neurones postganglionnaires parasympathiques sont cholinergiques.
- Les neurones postganglionnaires orthosympathiques sont noradrénergiques (sauf au niveau des glandes sudoripares et des vaisseaux cutanés qui sont cholinergiques).
-
Système parasympathique :
-
-
- Neurotransmetteur : Acétylcholine
- Synthèse :
- Neurotransmetteur : Acétylcholine
-
Au niveau de la terminaison synaptique, à partir d’acétylCoA et de choline. La réaction est catalysée par la choline acétylTransférase ou CHAT.
- Stockage :
Initialement synthétisées au niveau du soma, des vésicules issues du réseau trans-golgien sont acheminés grâce aux microtubules vers le bouton synaptique pour y être remplies d’acétylcholine. Ces vésicules sont organisées en deux pools : l’un de réserve, et l’autre prêt à l’exocytose dans la zone active.
- Libération :
L’arrivée du potentiel d’action dans le bouton synaptique induit l’ouverture des canaux calciques voltage dépendants. Le Ca2+ pénètre dans le MIC grâce au gradient électrochimique provoquant la fusion chimique des vésicules à la membrane présynaptique. L’excès de Ca2+ est éliminé grâce aux pompes calciques ATP-dépendantes.
- Elimination :
Après avoir exercé ses effets, l’Ach est hydrolysée par l’acétylcholinestérase en acide acétique et en choline. La choline est recaptée par la membrane présynaptique grâce à un mécanisme de transport actif.
- Récepteurs :
Une fois libérée dans la fente synaptique, l’Ach se fixe sur ses récepteurs spécifiques. Concernant le système parasympathique, ils sont muscariniques.
Il existe plusieurs types de récepteurs muscariniques : M1 (sécrétion acide de l’estomac), M2 (ralentissement du cœur ou encore innervation des glandes salivaires et lacrymales…). D’autres types M3, M4, M5 existent, mais leur fonction est moins connue.
- Pharmacologie :
Les agents qui agissent comme l’acétylcholine sont appelés parasympathomimétiques. Ils activent les récepteurs muscariniques. L’on cite comme exemple la muscarine ou la pilocarpine qui sont dits agonistes muscariniques.
Il existe des substances qui potentialisent l’action de l’Ach en inhibant l’action hydolytique de l’acétylcholinestérase. Ces agent (la néostigmine, la physostigmine, la DFP…) augmentent ainsi la quantité d’Ach disponible au niveau de la fente synaptique.
Ces récepteurs sont bloqués par des antagonistes muscariniques tels que l’atropine et la scopolamine.
Système sympathique :
-
- Neurotransmetteurs : Noradrénaline, adrénaline
- Synthèse :
- Neurotransmetteurs : Noradrénaline, adrénaline
Elle s’effectue à partir de l’acide aminé essentiel la tyrosine qui rejoint la cellule par un transporteur spécifique.
Tyrosine à tyrosine hydroxylase à DOPA à DOPA décarboxylase à dopamine à dopamine-β- hydroxylase à noradrénaline à phényléthanolamine-N-méthylTransférase à adrénaline.
- Stockage et libération :
La noradrénaline est stockée aux extrémités des fibres sympathiques postganglionnaires dans des granules. Ces dernières sont synthétisées dans le soma du neurone puis migrent le long de l’axone vers le bouton synaptique.
La NA stimule les récepteurs postsynaptiques alpha et béta.
- Dégradation :
Après avoir exercé son action, elle a deux destinées possibles : elle peut soit être recaptée par la membrane présynaptique pour être dégradée par la monoamine oxydase, soit diffuser dans la fente synaptique et y être dégradée.
- Récepteurs :
Il existe deux types de récepteurs adrénergiques : les récepteurs α et les récepteurs β, eux-mêmes subdivisés en deux familles chacun : α1, α2, β1 et β2.
Certains récepteurs ne sont présents que sur la membrane postsynaptique comme les récepteurs α1, d’autres sont présents sur les membranes pré et postsynaptiques comme les récepteurs α2.
La noradrénaline se fixe en priorité sur les récepteurs de type α, tandis que l’adrénaline active les récepteurs α et β de la même manière.
La relative richesse des organes cibles en récepteurs membranaires alpha et bêta définit leur affinité à la NA ou à l’adrénaline.

– Pharmacologie :
Les agents qui agissent comme la noradrénaline sont appelés sympathomimétiques. Ils activent
les récepteurs ardénergiques. L’on cite comme exemple l’adrénaline ou la méthoxamine qui sont dits agonistes adrénergiques.
Il existe des substances qui agissent sur une seule famille ou sous-famille de récepteurs seulement. C’est le cas de la phénylnéphrine (α), l’isoprénaline (β) et du salbutamol (β2).
Il existe des substances qui agissent sur la fibre présynaptique en provoquant la sécrétion des granules de noradrénaline. C’est le cas de l’éphédrine, la thyramine et l’amphétamine. Ils sont considérés comme sympathomimétiques indirects. A noter que la cocaïne et l’amphétamine inhibent la recapture de la noradrénaline et de la dopamine.
Ces récepteurs sont bloqués par des antagonistes adrénergiques tels que la phénoxybenzamine et la phentolamine (α), la prazosine (α1), la yohimbine (α2) et le propanolol (β).
D-Centres de contrôle du SNA :
Le SNA est contrôlé par des noyaux du tronc cérébral, lui-même modulés par d’autres structures : l’hypothalamus, la substance grise périaqueducale, le système limbique, cortex préfrontal…
E-Afférences viscérales :
Le neurone ganglionnaire spinal de la corne dorsale de la moelle épinière envoie une branche axonique qui se termine au niveau des récepteurs viscéraux et une seconde branche qui se termine dans la partie superficielle de la corne dorsale de la moelle où elle sera reliée à des neurones sympathiques et parasympathiques du SNA situés dans le noyau intermédiolatéral. L’arc réflexe est ainsi bouclé.
F-Mise en jeu du SNA :
Réflexes :
Réflexes végétatifs cardiovasculaires :
L’augmentation de la pression artérielle stimule les barorécepteurs des gros vaisseaux. Les fibres afférentes transmettent l’information au centre végétatif qui va inhiber le tonus sympathique exercé sur le cœur et les vaisseaux, provoquant ainsi une baisse de tension.
Réflexes végétatifs digestifs :
-
- La nourriture et son odeur déclenchent des influx nerveux cheminant vers les noyaux glossopharyngien et vagal du tronc cérébral. Ce dernier, par le biais des nerfs parasympathiques efférents, stimule les glandes sécrétrices de la partie haute du tube
- La distension du rectum provoque un réflexe parasympathique responsable des contractions péristaltiques du côlon aboutissant à la défécation.
Activité tonique :
Les nerfs sympathiques et parasympathiques envoient continuellement des influx nerveux (fréquence constante de PA) aux organes effecteurs. C’est ce que l’on appelle l’activité tonique de base des neurones préganglionnaires.
Elle permet en outre d’avoir une réponse ternaire aux stimuli de l’environnement.
Le principe de cette réponse ternaire est le suivant : dans les conditions physiologiques normales de pression artérielle, les fibres sympathiques envoient des PA à une fréquence donnée stable. En cas d’hypertension artérielle, cette fréquence va diminuer, et en cas d’hypotension, elle va augmenter. Cela implique l’existence de trois types de réponses traductibles par le centre végétatif. Ce phénomène augmente la capacité de mise en jeu physiologique de ces neurones.
Hypersensibilité à la dénervation :
En cas de section du nerf sympathique ou parasympathique, l’organe dénervé devient plus sensible à l’action de son neurotransmetteur. En effet, lorsqu’un organe se trouve dénervé, il a tendance à produire plus de récepteurs membranaires et devient ainsi beaucoup plus propice à être stimulé par le neuromédiateur qui l’active.
G-Effets du SNA sur les effecteurs viscéraux :
Œil :
Sympathique :
Ganglion cervical supérieur à stimulation du nerf sympathique à sécrétion de NA à fixation du NA sur les récepteurs α à contraction des muscles radiaires de l’iris à dilatation de la pupille (mydriase).
Parasympathique :
Ganglion ciliaire à stimulation nerf parasympathique (III) à sécrétion d’Ach à fixation de l’Ach sur les récepteurs muscariniques à contraction du sphincter pupillaire à constriction de la pupille (myosis).
Glandes exocrines :
Sympathique :
Le système sympathique contrôle les glandes sudoripares. En les stimulant, il induit une forte sudation. Cette innervation sympathique constitue toutefois une exception à la règle puisqu’elle est cholinergique.
Parasympathique :
La stimulation parasympathique induit une forte sécrétion des glandes lacrymales, nasales et salivaires riche en eau, sels minéraux et amylase.
Tube digestif :
Sympathique :
Une forte stimulation sympathique inhibe le péristaltisme intestinal, contracte les sphincters et induit donc un ralentissement de la progression du bol alimentaire dans l’intestin.
Parasympathique :
Le plexus d’Auerbach (véritable système nerveux autonome composé de plexus sensitivomoteurs) est sensible à l’innervation parasympathique en stimulant le péristaltisme et relâchant les sphincters. Ceci induit une accélération de la progression du bol alimentaire.
La défécation, comme la miction, résulte d’un réflexe parasympathique. Cependant, elle est toujours sous le contrôle de la volonté grâce au nerf pudendal interne.
Cœur et vaisseaux :
Sympathique :
-
- Augmente le rythme et la contraction.
- Contracte la plupart des vaisseaux (notamment ceux de la peau et des viscères).
- Tend à augmenter la pression artérielle (effets sur le cœur et la médullosurrénale).
Parasympathique :
-
- Diminue le rythme cardiaque et sa force de contraction.
- Sans effet sur les vaisseaux sauf au niveau de la face.
- Abaisse la pression artérielle.
Médullosurrénale :
La stimulation sympathique induit une sécrétion accrue d’adrénaline et de noradrénaline dans le sang. Elle joue un rôle majeur dans la régulation de la pression artérielle.
Cela va sans dire que l’effet de la médullosurrénale va au-delà que celui du système sympathique proprement dit. En effet, par le biais de ses sécrétions endocrines, elle agit sur le métabolisme de nombreuses cellules. Son action hormonale est également prolongée. Cependant, leurs actions sont similaires.
La médullosurrénale et le système sympathique se potentialisent tant qu’ils agissent ensemble simultanément.
Vessie :
Sympathique :
-
- Inhibition tonique du détrusor (musculeuse de la vessie) par les récepteurs β.
- Excitation du trigone vésical et du sphincter interne par les récepteurs α.
- Empêche la vidange de la vessie alors qu’elle est vide.
Parasympathique :
-
- Stimulation du détrusor.
- Inhibition du sphincter.
- Entraine la miction.
Le sphincter externe de l’uretère reçoit également une innervation motrice du système nerveux somatique, relevant donc du contrôle volontaire.
La miction est commandée par un réflexe végétatif contrôlé par la moelle ainsi que par la volonté. Ces mécanismes réflexes sont actifs chez le nouveau-né, puis sont peu à peu remplacés par le contrôle supraspinal.
Ce circuit réflexe met en jeu des mécanorécepteurs à la distension de la paroi vésicale. Lorsque la tension est importante, la miction est déclenchée tant par l’inhibition des projections sympathiques, que par l’activation des parasympathiques. La décision revient toutefois à la volonté, qui laissera s’exprimer le réflexe ou le réprimera. Le mécanisme est identique pour la défécation.
D’une manière générale :
Le système sympathique inhibe la motricité des voies biliaires, des uretères et des bronches, tandis que le parasympathique les stimule.
Le système sympathique augmente le métabolisme de base et stimule la glycogénolyse hépatique et musculaire.